 2015, Vol. 20
2015, Vol. 20



2 中国科学院大学, 北京100049
2 University of Chinese Academy of Sciences, Beijing 100049
植被是陆地生物圈的主体之一,它通过光合、呼吸、蒸腾等过程影响陆面和大气之间物质和能量的交换,在调节全球碳平衡、减缓大气中二氧化碳(CO2)等温室气体浓度上升以及维护全球气候稳定等方面具有不可替代的作用(朴世龙等,2001)。单位时间内植被通过光合作用所固定有机碳的量称为植被总初级生产力(Gross Primary Productivity,GPP),又称总第一性生产力,它决定了进入陆地生态系统的初始物质和能量,是植被生长的物质和能量来源,影响植被各碳库的分配量、净初级生产力(Net Primary Productivity,NPP),以及净生态系统交换(Net Ecosystem Exchange,NEE)等;另一方面,GPP受光照强度、大气CO2浓度、温度、水分、矿物质元素(如氮、磷等)的影响。因此GPP是联系陆地碳循环与大气碳库的重要纽带,在整个陆地生态系统碳循环中起到了至关重要的作用,引起了生态学家、碳循环与气候变化领域专家的广泛关注(Zhao et al.,2005;Yuan et al.,2007;王鹤松等,2010;Mao et al.,2012)。
目前,大尺度植被生产力的研究主要依赖于模型模拟,其大致可分为气候生产力模型、光能利用率模型以及生态生理过程模型。通过建立温度、降水、蒸散量等气候因子与植被干物质的关系来估算生产力(通常是植被潜在生产力)的气候生产力模型,以Miami模型、Thornthwaite纪念模型、Chikugo模型(Lieth,1975;Uchijima and Seino,1985)为代表。光能利用率模型考虑限制植物生长的资源(如水、氮、光照等)对生产力的影响,根据光合有效辐射(PAR)和最大光能利用率推求植被潜在生产力,并将这些限制因子与生产力的关系通过转换因子相联系,计算实际植被生产力。这类模型主要包括CASA模型(Potter et al.,1993)、GLO_PEM模型(Prince and Goward,1995;Goetz et al.,1999)、MODIS17算法(Zhao et al.,2005)、EC_LUE模型(Yuan et al.,2007)等,其依赖于实时遥感资料。基于植物生长发育和个体水平动态的生理生态学模型和基于生态系统内部功能过程的仿真模型,容易实现与气候(地球)系统模式的耦合以预测全球变化对植被生产力的影响以及土地覆盖分布的变化对气候的反馈作用(朱文泉等,2005),已成为生产力生态学研究的热点。这类模型主要有:LPJ模型(Sitch et al.,2003)、AVIM2模型(Ji,1995;Ji and Yu,1999;Lu and Ji,2002,2006)、CEASV模型(Cao and Woodward.,1998a,1998b)、BIOME- BGC模型(Running and Hunt,1993)、TEM模型(McGuire et al.,1995,1997)、CENTURY模型(Parton et al.,1993)等。
美国国家大气科学研究中心(NCAR)发展的陆面过程模式CLM4(Community L and Model version 4),包含基于生态机理的生物地球化学模块,可对植被、凋落物和土壤有机质中的碳、氮变量进行模拟,同时保持植被—雪—土壤间的水分和能量平衡预报(Oleson et al.,2010)。该模式所使用的驱动数据以及基于生态生理过程的模型结构和参数化可能会给模拟结果带来很大的不确定性(Zaehle et al.,2005;Jung et al.,2007)。Mao et al.(2012)利用2000~2009年MODIS(MODerate-resolution Imaging Spectroradiometer)卫星的GPP遥感反演产品对CLM4全球GPP模拟能力进行了评估,结果表明CLM4能较好刻画出全球GPP的空间分布格局,但对热带常绿林存在严重高估。为了进一步分析和评估其在区域尺度的模拟能力,本研究使用时间段相对较长的基于站点观测的升尺度数据MTE_ GPP(Jung et al.,2009),从空间分布、季节变化、年际变化等方面对CLM4模拟的中国区域1982~2004年GPP结果进行比较分析,以评估CLM4对中国区域GPP时空变化特征及规律的模拟能力,并探讨了不同土地覆盖资料对GPP模拟的影响。
2 模型、数据与试验设计本节将对本研究所采用的模型、输入数据(包括大气强迫数据和土地覆盖资料),以及用于结果评估的GPP通量站点观测和MTE_GPP进行介绍。
2.1 陆面过程模式CLM4陆面过程模式CLM4是气候系统模式CCSM4.0(Community Climate System Model,verion 4.0)和地球系统模式CESM1.0(Community Earth System Model,vesion1.0)中的陆面过程模式分量。与上一版本模型CLM3.5相比在水文过程、雪过程、有机土壤的水热性质等参数化方面均有很大改进。CLM4考虑了网格尺度内地表特征的差异、不同植被类型生态学变异以及不同土壤层水动力学和热力学上的差异性,并通过嵌套次网格实现。它包含生物地球物理过程、水文过程和生物地球化学过程参数化。对生物地球化学过程的描述包含3种模块:基于静态植被的光能利用率模型CASA,基于静态植被考虑碳和氮相互作用的CN模块以及基于动态植被(DGVM)的CN模块CNDV(分别记为CLM4- CASA、CLM4-CN、CLM4-CNDV)。在下面的模拟试验中我们使用CLM4-CN,它是基于陆地生物地球化学模型Biome-BGC(version 4.1.2)发展而来,预报变量包括陆地植被、凋落物和土壤有机质中的碳、氮以及植被—雪—土壤间的水和能量等。由于阳叶、阴叶属性的不同,其辐射传输、气孔导度及光合作用也不相同,因此CLM4-CN中GPP的模拟采用两叶模型,计算冠层阳叶与阴叶的光合作用之和,它受温度、气孔导度、土壤湿度、根分布以及土壤和叶片中氮含量等的影响(Oleson et al.,2010)。
2.2 数据 2.2.1 大气强迫数据Qian et al.(2006)基于NCAR/NCEP的1948~2004年大气数据集发展了水平分辨率约为1.875°(纬度)×1.875°(经度)的气象强迫场数据(以下简称该强迫数据为QIAN),它包括3 h风场、气温、比湿、气压场和6 h辐射、降水数据。温度与降水由月观测值与NCAR/NCEP再分析资料的月内变化相结合而得到;向下到达地表的太阳辐射通过站点观测的云覆盖异常对再分析资料变化和趋势进行校正并利用卫星资料对平均偏差进行校正;比湿通过已校正的温度与相对湿度对再分析比湿进行校正;风场和压力数据由6 h再分析资料直接插值而得。
2.2.2 土地覆盖数据精确的土地覆盖资料是研究陆地能量交换、水循环和生物地球化学过程的重要基础。CLM4中所用土地覆盖资料为Lawrence and Chase(2007)基于MODIS卫星遥感资料发展的地表覆盖数据集(以下将该数据简称为MODIS),空间分辨率为0.5°(纬度)×0.5°(经度),其将植被按叶、茎的光学性质,根分布参数和光合参数等划分为16种植物功能型(Plant Functional Types,PFTs),依次为裸土(Bare)、温带针叶常绿林(NEM Tr)、寒带针叶常绿林(NEB Tr)、寒带针叶落叶林(NDB Tr)、热带阔叶落叶常绿林(BET Tr)、温带阔叶常绿林(BEM Tr)、热带阔叶落叶林(BDT Tr)、温带阔叶落叶林(BDM Tr)、寒带阔叶落叶林(BDB Tr)、温带阔叶常绿灌木(BE Sh)、温带阔叶落叶灌木(BDM Sh)、寒带阔叶落叶灌木(BDB Sh)、C3极地草地(C3 AR)、C3非极地草地(C3 NA)、C4草地(C4)、农作物(Crop)。MODIS土地覆盖资料虽然有较高的整体分类精度,但在局部存在明显的分类错误,不能很好的满足陆表过程模拟的需要(冉有华等,2009)。
为了提高中国土地覆盖资料的精度,Ran et al.(2012)基于1:1000000植被图、2000年1:100000 土地利用图、1:1000000 沼泽—湿地图、冰川图和2001年MODIS土地覆盖图(MODIS 2001)发展了一套多源数据融合的1 km分辨率的中国土地覆盖资料(Multi-source Integrated Chinese L and Cover,MICLCover),其与美国地质调查局为国际地圈—生物圈计划建立的全球土地覆盖数据集(IGBPDISCover)和MODIS土地覆盖相比有很大改进。为方便对比试验,我们将MICLCover数据升尺度为0.5°(纬度)×0.5°(经度)(网格面积加权平均,以下将MICLCover数据简称为MICL)。与MODIS土地覆盖数据相比,MICL在中国区域的冰川、湖泊、湿地分别增加了0.5%、1.6%、1.1%,植物功能型中裸土、针叶林、阔叶林分别减少了14.6%、3.6%、1.9%,灌木、草地、农作物分别增加了1.8%、9.9%、5.0%。图 1a、1b分别为MODIS和MICL资料中国陆地网格中覆盖率最大的植物功能型(即主要植物功能型)分布(梁妙玲和谢正辉,2006)。与MODIS相比,MICL土地覆盖资料的较大差异体现为西部裸土(Bare)的减少,青藏地区C3极地草地(C3 AR)和内蒙地区C3非极地草地(C3 NA)的增加。
 | 图 1 中国区域主要植物功能型分布:(a)MODIS;(b)MICL Fig. 1 The dominant plant functional types in China for(a)MODIS(MODerate-resolution Imaging Spectroradiometer) and (b)MICL(Multi-source Integrated Chinese L and cover) |
中国陆地生态系统通量观测研究网(http://www.cerndata.ac.cn[2013-09-20])集合了中国不同区域的通量网数据,为区域碳循环研究和模型验证提供观测数据。本研究使用其中6个通量站的GPP观测资料对模型进行验证,包括长白山站(CBS)、禹城站(YC)、千烟洲站(QYZ)、鼎湖山站(DHS)、西双版纳站(XSBN)和海北站(HB),其详细信息见表 1。
|
|
表 1 通量观测站点信息 Table 1 Informations of six China flux sites |
FLUXNET数据库(www.fluxdata.org [2013- 09-20])包含了全球CO2、水、能量的涡度相关观测,这为全球陆地碳水循环研究提供了基础。然而这些观测点分布不均,且主要集中在北半球中纬地区,不能覆盖全球。为了获得时空连续的全球GPP数据集,Jung et al.(2009)采用机器学习—模型树集合算法(Model Tree Ensemble,MTE),将全球涡度相关通量观测数据进行经验性升尺度,得到空间分辨率为0.5°(纬度)×0.5°(经度)全球1982~2011年的月GPP数据集(http://www.bgc-jena.mpg.de/geodb/projects/Data.php[2013-07-23])。由于QIAN大气强迫截止到2004年,为保持二者时间段的一致,选取中国区域1982~2004年的数据作为观测来与模拟值比较(以下将该数据简称为MTE_GPP)。
2.3 试验设计本研究选取CLM4-CN进行离线模拟试验,使用QIAN1972~2004年大气强迫场为驱动,大气CO2浓度(367 ppmv)和气溶胶沉降速率选用2000年水平,研究区域为中国(15°N~55°N,70°E~140°E),空间分辨率为0.5°(纬度)×0.5°(经度),模式积分步长为1800 s,模拟结果为月输出。
本次模拟设计了两组试验,第一组试验使用MODIS资料,记为CLM4_M,为获取模式平衡态,本研究首先使用1972~2004年的大气强迫循环驱动模式650年(Kluzek,2011)。使用此刻得到的模式状态作为初始场,模拟1982~2004年。为探讨CLM4-CN模拟的GPP对土地覆盖资料的敏感性,第二组试验使用MICL代替MODIS资料,其余均与CLM4_M试验相同,记为CLM4_C。两组试验模拟得到的GPP分别记为CLM4_GPP和CLM4_ MICL_GPP。
3 结果分析我们首先利用2003~2004年长白山站、禹城站、千烟洲站、鼎湖山站、西双版纳站和海北站6个通量站的GPP观测数据对CLM4_GPP进行验证,然后对其空间分布、季节变化和年际变化3个方面与MTE_GPP进行比较分析。最后通过比较CLM4_GPP与CLM4_ MICL_GPP的差异探讨不同土地覆盖资料对GPP模拟的影响。
3.1 站点验证图 2给出了6个通量站2003~2004年GPP月观测值、MTE_GPP、CLM4_GPP和CLM4_ MICL_GPP的季节变化。可以看出,6个站点的MTE_GPP与观测值相关性均较好,量值也较为接近(表 2),表明MTE_GPP具有较高的准确性,可以作为观测用于3.2−3.5节中与CLM4_GPP比较。6个站点的CLM4_GPP与观测值季节变化一致,其中长白山站、千烟洲站、鼎湖山站、西双版纳站和海北站季节变化特征相似,表现为单峰变化,而禹城站由于为一年两熟作物类型,GPP呈双峰变化。各个站点的CLM4_GPP与观测在量值上存在不同程度的偏差(表 2),其中地处亚热带的千烟洲站和鼎湖山站全年存在较大高估(均方根偏差RMSD>100 gC m−2 month−1),Chen et al.(2011)和Bonan et al.(2011)研究表明热带和亚热带地区GPP的高估主要是由CLM4不精确的参数化方案引起的,例如计算光合速率时,选取受暗反应中Rubisco酶总量和活性限制的同化速率、受光反应中光照条件对电子输送速率限制的同化速率以及受光合产物利用程度限制(C3)或受PEP羧化速率(C4)限制的同化速率中的最小值,这会造成光合速率的突变;计算冠层尺度光合作用参数时,没有考虑叶片生理特征的垂直变化(即最大光合速率由冠层顶向下衰减的特性),这也会导致热带、亚热带地区GPP偏高。另外,GPP与植物的光合作用有关,植被的光合作用又受到各种养分的限制,如氮、磷等;我国南部、东部地区受到氮—磷混合限制,且南部地区主要是磷限制(Wang et al.,2010);这些地区地势低且土壤湿度较大,磷养分较容易流失,且土壤主要呈酸性,磷元素容易被固定失效;而目前CLM4- CN仅考虑了氮循环对GPP的限制,却没有考虑其他养分(如磷)的限制,这可能也是导致GPP高估的重要原因。而长白山站、西双版纳站和海北站偏差相对较小(RMSD<100 gC m−2 month−1)。长白山站在生长季模拟值存在正偏差而非生长季存在负偏差;禹城站在作物生长季存在低估;西双版纳站2003年全年和2004年下半年模拟值偏高,而2004年上半年模拟值则与观测非常吻合;海北站在生长季存在低估,非生长季存在高估。另外,对于长白山站、禹城站和海北站,CLM4_GPP表现出的植被生长季均要长于观测。
 | 图 2 6个通量站MTE_GPP、CLM4_GPP、CLM4_MICL_GPP与观测数据的比较:(a)长白山站;(b)禹城站;(c)千烟洲站;(d)鼎湖山站;(e)西双版纳站;(f)海北站 Fig. 2 Comparison of monthly MTE(Model Tree Ensemble)_GPP(Gross Primary Productivity),CLM4(Community L and Model version 4)_GPP,CLM4_MICL(Multi-source Integrated Chinese L and )_GPP with corresponding tower-based observations at six flux tower sites ofn China from 2003 to 2004:(a)Changbaishan;(b)Yucheng;(c)Qianyanzhou;(d)Dinghushan;(e)Xishuangbanna;(f)Haibei |
|
|
表 2 6个通量站CLM4_GPP、MTE_GPP、CLM4_MICL_ GPP与观测值的相关系数和均方根偏差 Table 2 Pearson correlations and RMSDs(Root-Mean-Square Differences)between CLM4_GPP,MTE_GPP,CLM4_ MICL_GPP,and observed GPP values at six flux tower sites of China |
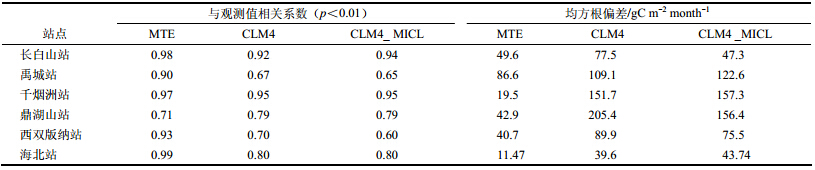
总体而言,CLM4-CN能较好地模拟以上6个站点GPP的季节变化,但由于模型模拟与观测样地尺度的差别、模型输入数据(大气强迫和土地覆盖)的不确定性以及模型参数化方案等原因,CLM4_GPP在量值上相比观测存在不同程度的偏差。
3.2 空间分布
基于MODIS17算法,目前MODIS陆地工作组已发布了2000~2012年的全球GPP产品(记为MODIS_GPP)。此处除MTE_GPP外,同时选取2000~2004年MODIS_GPP与模拟结果进行比较。图 3给出的是2000~2004年多年平均GPP空间分布。由图 3a可看出,在中国区域,模拟的CLM4_GPP整体分布格局表现为由东南向西北递减,其主要原因是我国西北地区植被稀少,气候干旱,而东部地区气候湿润,水热条件较好,能更好地满足植被生长需求(陶波等,2003),这一分布格局与中国区域降水分布颇为相似(何勇等,2006)。东南部由于处于热带雨林区,如广东、广西、台湾等地区GPP值较高,可达3500~4000 gC m−2 a−1;而在西藏、青海、甘肃、内蒙、新疆等地的部分地区,GPP值较小,低于500 gC m−2 a−1。图 3b、3c中空白区分别为MTE_GPP和MODIS_GPP数据缺省区域,二者亦呈现东南向西北减小的空间分布格局,与CLM4_GPP的空间相关系数分别为0.93、0.87(均通过显著性检验,p<0.01),但在量值上有差别。整个中国区域由MTE_GPP和MODIS_GPP计算得到的年GPP量分别为6.9 PgC a−1和5.9 PgC a−1,而CLM4_M模拟得到的年GPP量为13.7 PgC a−1,几乎是MTE_GPP和MODIS_GPP的2倍。由图 3d、3e可看出,CLM4_M模拟的GPP在中国普遍偏高。对于CLM4_M模拟结果较大的地区(即低纬地区)正偏差均较大,高于900 gC m−2 a−1;在内蒙、青海、西藏等地区,正偏差较小,仅不足300 gC m−2 a−1;而在中高纬地区如内蒙、甘肃、新疆等地,表现为负偏差。总体而言,对于中国区域,CLM4_M模 拟的GPP在低纬地区和东北正偏差较大,而在其他地区,如内蒙古、青海、新疆等西部地区,偏差较小。
 | 图 3 2000~2004年多年平均GPP分布:(a)CLM4_GPP;(b)MTE_GPP;(c)MODIS_GPP;(d)CLM4_GPP与MTE_GPP的差值;(e)CLM4_GPP与MODIS_GPP的差值 Fig. 3 Means of yearly GPP in China over the period from 2000 to 2004 for(a)CLM4_GPP,(b)MTE_GPP,(c)MODIS_GPP,(d)difference between CLM4_GPP and MTE_GPP,and (e)difference between CLM4_GPP and MODIS_GPP |
除了模型参数化方案对GPP模拟结果产生影响外,大气强迫场也具有重要影响,为此,我们选取位于亚热带的贵阳、南京、杭州和汕头4个地面气象观测站1982~2004年地表入射太阳辐射(FSDS)、降水(PREC)、比湿(QBOT)和大气温度(TBOT)的月均值,并将这些观测值与QIAN大气强迫数据进行对比(图 4)。与观测相比,QIAN地表入射太阳辐射、降水、比湿和大气温度存在不同程度的偏差,其中大气温度(图 4d)与观测非常吻合(相关系数r=0.99),且偏差较小(平均偏差MBE=0.23 °C);降水(图 4b)分布较为分散(RMSE=1.53 mm d−1);而比湿(图 4c)和地表入射太阳辐射(图 4a)整体存在正偏差。由于植被的光合作用一般会随地表入射太阳辐射的增加而增强(Mercado et al.,2009),因此,QIAN地表入射太阳辐射的高估很可能是造成该区域GPP高估的另一重要原因。不仅如此,太阳辐射变化对生态系统光合作用的影响还取决于散射辐射比例的变化以及阳叶、阴叶在冠层中的比例(李登秋等,2014),而目前CLM4大气强迫场仅含有总辐射量(Qian et al.,2006;Oleson et al.,2010),这使得大气强迫数据(尤其是地表入射太阳辐射)对CLM4_GPP模拟的影响还需要更多的观测数据进行深入研究。
 | 图 4 1982~2004 年 QIAN 大气强迫数据与地面站点观测的比较:( a)地表入射太阳辐射( FSDS);( b)降水( PREC);( c)大气比湿( QBOT);( d)大气温度( TBOT)。 MBE 表示平均偏差, RMSE 表示均方根误差, r 表示相关系数 Fig. 4 Comparison of monthly surface downward solar radiation (FSDS), precipitation (PREC), atmospheric specific humidity (QBOT), and atmospheric air temperature (TBOT) between QIAN on 0.5° grid with in situ measurements at Guiyang, Nanjing, Hangzhou, and Shantou sites from 1 Jan 1982 to 31 Dec 2004. MBE indicates mean bias, RMSE indicates root-mean-square error, and r indicates correlation coefficient |
图 5给出的是CLM4_GPP与MTE_GPP多年平均季节差异的空间分布。CLM4-CN模拟结果都以高估为主,尤其是春秋季节。夏季(图 5c),有少部分区域出现―60 gC m−2 month−1左右的低估。在冬季(图 5a),高估区域范围缩小,偏差值也减小。对于低纬30°N以南地区,四季均存在高估。
 | 图 5 CLM4_GPP与MTE_GPP季节差异分布:(a)冬季;(b)春季;(c)夏季;(d)秋季 Fig. 5 The spatial differences of annual mean GPP during 1982 and 2004 between CLM4_GPP and MTE_GPP in (a) winter (DJF), (b) spring (MAM), (c) summer (JJA), and (d) autumn (SON) |
图 6给出了多年平均下我国13种主要植物功能型CLM4_GPP与MTE_GPP的年内循环(此处为了统一,CLM4_GPP与MTE_GPP均按MODIS中国主要植物功能型划分)。CLM4_GPP和MTE_GPP各植物功能型的季节特征相似,表现为单峰变化,GPP在春季随植被生长而快速上升,在6月末8月初达到最大值,在秋季由于落叶(落叶林)、气温下降等原因迅速减小,到冬季降到最低。表 3中各植物功能型CLM4_GPP和MTE_GPP年内循环序列相关系数均大于0.9(通过显著性检验 p<0.01),二者年内循环具有较好的一致性。
 | 图 6 中国主要植物功能型CLM4_GPP和MTE_GPP年内循环 Fig. 6 The annual cycles of CLM4_GPP and MTE_GPP of dominant PFTs(Plant Functional Types)in China |
|
|
表 3 中国主要植物功能型CLM4_GPP与MTE_GPP年内循环的相关系数、均方根偏差、年际时间序列的相关系数、年际变率、变化趋势 Table 3 Pearson correlations between intra-annual and inter-annual CLM4_GPP and MTE_GPP time series of dominant PFTs in China,RMSDs,interannual variabilities,and the trends |
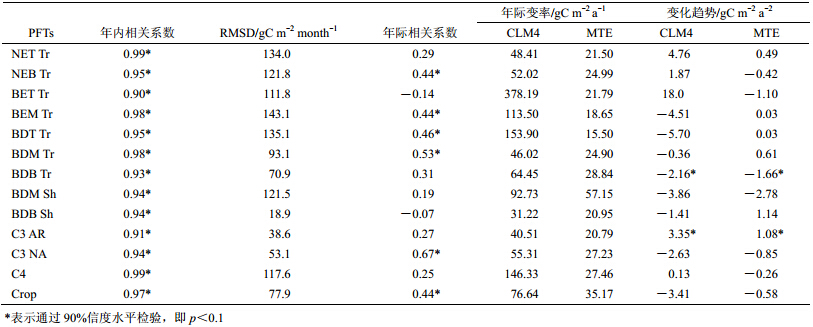
在量值上,对于温带阔叶落叶林(图 6f)、寒带阔叶落叶林(图 6g)、寒带阔叶落叶灌木(图 6i)、C3极地草地(图 6j)、C3非极地草地(图 6k)和农作物(图 6m),CLM4_GPP与MTE_GPP较为接近(RMSD<100 gC m−2 month−1)(表 3),其中寒带阔叶落叶林、寒带阔叶落叶灌木、C3极地草地分别在4~11月、6~9月、4~12月存在10~20 gC m−2 month−1的高估;且寒带阔叶落叶灌木在10~11月间存在微小低估;而温带阔叶落叶林、C3非极地草地和农作物全年分别存在40~80、10~20、40~60 gC m−2 month−1的高估。其他各植物功能型GPP模拟值相对于MTE_GPP,全年存在较大正偏差(RMSD>100 gC m−2 month−1)。同时,对于温带针叶常绿林(图 6a)、寒带针叶常绿林(图 6b)、温带阔叶落叶林(图 6f)、寒带阔叶落叶林(图 6g)、C3极地草地(图 6j)和C3非极地草地(图 6k)而言,CLM4表现出的生长季均长于MTE_GPP。
3.4 年际特征图 7中给出了中国主要植物功能型1982~2004年CLM4_GPP和MTE_GPP的年时间序列。对于13种植物功能型,CLM4_GPP表现出的年际变率均大于MTE_GPP(表 2),其中热带阔叶落叶常绿林、温带阔叶常绿林、热带阔叶落叶林和C4草地年际变化较大,超过100 gC m−2 a−1。寒带针叶常绿林、温带阔叶常绿林、温带阔叶落叶林、热带阔叶落叶林、C3非极地草地、农作物的CLM4_GPP与MTE_GPP存在很好的年际对应关系。热带针叶常绿林、寒带阔叶落叶林和C3极地草地CLM4_GPP与MTE_GPP变化趋势表现一致(表 2)且通过显著性检验(p<0.1),其中寒带阔叶落叶林分别以―2.16和―1.16 gC m−2 a−2的速率减小;而热带针叶常绿林、C3极地草地的CLM4_GPP和MTE_GPP分别以4.76、0.49、3.35、1.08 gC m−2 a−2的速率增加。对于其他植物功能型,除温带阔叶落叶灌木、C3非极地草地和农作物外,大多表现为相反的变化趋势。
 | 图 7 1982~2004年中国主要植物功能型CLM4_GPP与MTE_GPP 年际特征 Fig. 7 The time series of CLM4_GPP and MTE_GPP of dominant PFTs in China from 1982 to 2004 |
气候因素(包括太阳辐射、大气温度、降水等)、大气CO2浓度、土壤(土壤类型、理化性状和肥力等)、生物(植物种类及其生物学性状等)和人类活动(毁林开荒、植树造林等)都会影响植物的光合作用、呼吸作用等过程,影响陆地生态系统碳循环。其中,在气候因子中太阳辐射、温度、降水是影响陆地生态系统碳循环的3个重要因素。植被对气候变化响应的滞后时间一般在月或季节时间尺度上(朱文泉等,2007),并不会掩盖其年际特征,因此主要在年际尺度上探讨气候因子与GPP的关系。
由表 4可看出,在研究时段内,整个中国区域GPP总量年际变化(r=0.76,p<0.1)与降水量显著正相关,而与年平均温度(r=0.28,p>0.1)之间的相关关系并不显著。因此降水是控制全国GPP总量年际变化的主要气候因子。这与陶波等(2003)、何勇等(2006)、黄珏等(2013)研究结果一致。同时这也可能是本文3.2节中GPP空间分布表现与降水格局一致的原因。
|
|
表 4 整个中国区域GPP总量年际变化与降水、辐射、温度的相关关系 Table 4 Pearson correlations between the spatial averaged annual GPP and climate variables |
根据地理位置将中国分为9个子区域(Liu et al.,2013),如图 8a所示,其中:Ⅰ为东北地区(黑龙江、吉林、辽宁)、Ⅱ为内蒙古地区、Ⅲ为西北地区(甘肃、宁夏、新疆)、Ⅳ为华北地区(北京、河北、河南、山东、山西、陕西、天津)、Ⅴ为华中地区(湖北、湖南)、Ⅵ为青海—西藏高原地区(青海、西藏)、Ⅶ为东南地区(安徽、福建、江苏、江西、上海、台湾、浙江)、Ⅷ为华南地区(广东、广西、海南、香港、澳门)、Ⅸ为西南地区(贵州、四川、云南、重庆)。各个子区域的CLM4_GPP与MTE_GPP的多年平均值如图 8b所示。由图 8b可见,华南地区GPP值最高,其次是东南、华中和西南地区,这些区域的主要植物功能型为农作物和温带针叶常绿林(图 1a);西北地区以裸土为主GPP值最小。从中国9个子区域GPP的高低排序来看,CLM4模拟得到GPP格局与MTE_GPP一致,但在量值上除西北地区,所有区域CLM4_GPP都存在正偏差,尤其是在华中、青海—西藏、东南、华南和西南地区,CLM4_GPP约为MTE_GPP的2倍,东 北、内蒙和华北地区偏差相对较小,对于西北地区二者数值相当。
 | 图 8 (a)中国九大地理分区和(b)9个子区域多年平均下的CLM4_GPP与MTE_GPP Fig. 8 (a)The nine sub-regions of China and (b)means of yearly CLM4_GPP and MTE_GPP in the nine sub-regions from 1982 to 2004 |
降水和温度对GPP的影响在不同地区存在较大的差异,其中东北(r=0.65,p<0.01)、内蒙(r=0.84,p<0.01)、西北(r=0.61,p<0.01)、华北(r=0.73,p<0.01)地区,由于降水匮乏,年降水量低于700 mm,尤其是西北地区年降水量低于200 mm,因此降水是控制其GPP的主要因子。而对于华中(r=0.73,p<0.01)和华南(r=0.35,p<0.01)地区年均温度的影响更为重要,这与陶波等(2003)研究结果一致,但与黄珏等(2013)研究结果并不一致,这可能与模式自身的参数化过程有关,具体原因值得进一步探讨。
3.6 不同土地覆盖资料对GPP的影响图 9分别给出了多年平均CLM4_MICL_GPP及与CLM4_GPP差值的空间分布。由图 9a可以看出,CLM4_MICL_GPP与CLM4_GPP分布特征相似,表现为由东南向西北递减。全国CLM4_MICL_ GPP多年平均值为13.6 PgC a−1,低于CLM4_GPP。中国大部分区域由不同土地覆盖资料造成的GPP变化显著(图 9b),其中东北、西南、华北和内蒙部分地区降幅超过20%,而西北地区、青藏、西北、内蒙和华北部分地区由于裸土的减少,增幅超过20%,有的区域甚至超过40%。
 | 图 9 多年平均(a)CLM4_MICL_GPP和(b)CLM4_MICL_GPP相对CLM4_GPP变化百分比的空间分布(网格阴影区为t检验通过95%信度水平区域) Fig. 9 Spatial distributions of(a)mean CLM4_MICL_GPP and (b)the difference percentage between mean CLM4_MICL_GPP and CLM4_GPP(grid shadow regions: over 95% confidence level) |
与CLM4_GPP相比(图 2),长白山站、鼎湖山站和西双版纳这3个通量站CLM4_MICL_GPP与观测的偏差变小,二者更为接近(表 2)。而这3个站点在MICL资料中的主要植物功能型分别为寒带阔叶落叶林、热带阔叶落叶林和热带阔叶落叶林,相比MODIS资料中的寒带针叶常绿林、温带针叶常绿林和温带阔叶常绿林,虽与站点实际植被类型并不完全一致(表 1),但更为接近。由于观测样地尺度和模式模拟尺度的不匹配,其他3个站点的植被类型在其对应模式网格中并不具有代表性,因此CLM4_MICL_GPP并未得到改善。因此,土地覆盖资料的不准确很可能是造成CLM4_GPP偏差的另一重要原因。
4 结论本文通过将CLM4模拟的中国区域1982~2004年GPP与基于观测的升尺度数据MTE_GPP进行比较,评估了CLM4对中国区域GPP时空变化特征及规律的模拟能力,并探讨了不同土地覆盖资料对GPP模拟的影响。结果表明:CLM4-CN能够较好刻画中国区域GPP空间分布格局和各植物功能型的GPP季节特征,但量值存在偏差;CLM4_GPP不同植物功能型的年际变率均大于MTE_GPP,仅热带针叶常绿林、寒带阔叶落叶林和C3极地草地的CLM4_GPP与MTE_GPP变化趋势一致;降水是研究时段内控制整个中国区域GPP的主要气候因子;土地覆盖资料的不同会造成GPP的显著变化,精确的土地覆盖数据可以减小模型GPP模拟的不确定性。
磷循环是生态系统中另一必不可少的物质循环,如何在CLM中实现磷循环动态变化以及碳—氮—磷的相互作用过程,对改善CLM4_GPP模拟具有重要作用。同时计算叶光合速率时,应避免其取值的突变;计算冠层尺度光合作用参数时,考虑叶片生理特征的垂直变化特性将会大大减小CLM4对GPP的高估。
另外,本研究中采用的QIAN大气强迫场存在较大的不确定性,由此造成的误差不可避免。试验中采用的是单一大气CO2浓度值,未考虑其空间差异与时间变异,由此造成的误差也是不可忽略的。因此,在未来的工作中我们将尽可能的从以上方面完善模式及其模拟以改善CLM4-CN的GPP模拟,从而减小陆地生态系统碳循环的不确定性。
致谢 感谢中国陆地生态系统通量观测研究网(http://www.cerndata.ac.cn[2013-09-20])共享长白山、禹城、千烟洲、鼎湖山、西双版纳和海北6个通量站点的相关通量观测资料。感谢审稿专家提出的宝贵意见。| [1] | Bonan G B, Lawrence P J, Oleson K W, et al. 2011. Improving canopy processes in the community land model version 4 (CLM4) using global flux fields empirically inferred from FLUXNET data [J]. J. Geophys. Res., 116, G02014, doi: 10.1029/2010JG001593. |
| [2] | Cao M K, Woodward F I. 1998a. Dynamic responses of terrestrial ecosystem carbon cycling to global climate change [J]. Nature, 393: 249-252. |
| [3] | Cao M K, Woodward F I. 1998b. Net primary and ecosystem production and carbon stocks of terrestrial ecosystems and their responses to climate change [J]. Global Change Biology, 4: 185-198. |
| [4] | Chen H S, Dickinson R E, Dai Y J, et al. 2011. Sensitivity of simulated terrestrial carbon assimilation and canopy transpiration to different stomatal conductance and carbon assimilation schemes [J]. Climate Dyn., 36: 1037-1054. |
| [5] | Goetz S J, Prince S D, Goward S N, et al. 1999. Satellite remote sensing of primary production: An improved production efficiency modeling approach [J]. Ecological Modelling, 122: 239-255. |
| [6] | 何勇, 姜允迪, 丹利, 等. 2006. 中国气候、陆地生态系统碳循环研究[M]. 北京: 气象出版社, 66-67. He Yong, Jiang Yundi, Dan Li, et al. 2006. Research of Climate and Terrestrial Ecosystem Carbon Cycle of China (in Chinese) [M]. Beijing: China Meteorological Press, 66-67. |
| [7] | 黄珏, 陈海山, 俞淼. 2013. 1981~2008年中国陆地植被NPP对气候变化响应的敏感性试验 [J]. 大气科学学报, 36 (3): 316-322. Huang Jue, Chen Haishan, Yu Miao. 2013. Sensitivity experiments on response of terrestrial net primary productivity in China to climate change during 1981-2008 [J]. Transactions of Atmospheric Sciences (in Chinese), 36 (3): 316-322. |
| [8] | Ji J J. 1995. A climate-vegetation interaction model: Simulating physical and biological processes at the surface [J]. Journal of Biogeography, 22: 445-451. |
| [9] | Ji J J, Yu L. 1999. A simulation study of coupled feedback mechanism between physical and biogeochemical processes at the surface [J]. Chinese Journal of Atmospheric Sciences, 23 (4): 439-448. |
| [10] | Jung M, Vetter M, Herold M, et al. 2007. Uncertainties of modeling gross primary productivity over Europe: A systematic study on the effects of using different drivers and terrestrial biosphere models [J]. Global Biogeochemical Cycles, 21 (4), GB4021, doi: 10.1029/2006GB002915. |
| [11] | Jung M, Reichstein M, Bondeau A. 2009. Towards global empirical upscaling of FLUXNET eddy covariance observations: Validation of a model tree ensemble approach using a biosphere model [J]. Biogeosciences, 6: 2001-2013. |
| [12] | Kluzek E. 2011. CCSM research tools: CLM4.0 user's guide documentation. Available online at http://www.cesm.ucar.edu/models/cesm1.0/clm/ models/lnd/clm/doc/UsersGuide/book1.html. |
| [13] | Lawrence P J, Chase T N. 2007. Representing a new MODIS consistent land surface in the community land model (CLM 3.0) [J]. J. Geophys. Res., 112: G01023, doi: 10.1029/2006JG000168. |
| [14] | 李登秋, 周艳莲, 居为民, 等. 2014. 太阳辐射变化对亚热带人工常绿针叶林总初级生产力影响的模拟分析 [J]. 植物生态学报, 38 (3): 219-230. Li Dengqiu, Zhou Yanlian, Ju Weimin, et al. 2014. Modelling the effects of changes in solar radiation on gross primary production in subtropical evergreen needle-leaf plantations [J]. Chinese Journal of Plant Ecology (in Chinese), 38 (3): 219-230. |
| [15] | 梁妙玲, 谢正辉. 2006. 我国气候对植被分布和净初级生产力影响的数值模拟 [J]. 气候与环境研究, 11 (5): 582-592. Liang Miaoling, Xie Zhenghui. 2006. Simulations of climate effects on vegetation distribution and net primary production in China [J]. Climatic and Environment al Research (in Chinese), 11 (5): 582-592. |
| [16] | Lieth H. 1975. Modeling the primary productivity of the world [C]./Primary Productivity of the Biosphere. Berlin, Heidelberg: Springer, 237-263. |
| [17] | Liu Y B, Ju W M, He H L, et al. 2013. Changes of net primary productivity in China during recent 11 years detected using an ecological model driven by MODIS data [J]. Frontiers of Earth Science, 7 (1): 112-127. |
| [18] | Lu J H, Ji J J. 2002. A simulation study of atmosphere-vegetation interactions over the Tibetan Plateau. Part I: Physical fluxes and parameters [J]. Chinese Journal of Atmospheric Sciences, 26 (1): 111-126. |
| [19] | Lu J H, Ji J J. 2006. A simulation and mechanism analysis of long-term variations at land surface over arid/semi-arid area in North China [J]. J. Geophys. Res., 111 (D9): D09306. |
| [20] | Mao J F, Thornton P E, Shi X Y, et al. 2012. Remote sensing evaluation of CLM4 GPP for the period 2000-09 [J]. J. Climate, 25 (15): 5327-5342. |
| [21] | McGuire A D, Melillo J M, Kicklighter D W, et al. 1995. Equilibrium responses of soil carbon to climate change: Empirical and process-based estimates [J]. Journal of Biogeography, 22 (4-5): 785-796. |
| [22] | McGuire A D, Melillo J M, Kicklighter D W, et al. 1997. Equilibrium responses of global net primary production and carbon storage to doubled atmospheric carbon dioxide: Sensitivity to changes in vegetation nitrogen concentration [J]. Global Biogeochemical Cycles, 11 (2): 173-189. |
| [23] | Mercado L M, Bellouin N, Sitch S, et al. 2009. Impact of changes in diffuse radiation on the global land carbon sink [J]. Nature, 458: 1014- 1017. |
| [24] | Oleson K W, Lawrence D M, Bonan G B, et al. 2010. Technical description of version 4.0 of the community land model (CLM) [R]. NCAR Tech. Note NCAR/TN-4781STR, 257 pp. |
| [25] | Parton W J, Scurlock J M O, Ojima D S, et al. 1993. Observations and modeling of biomass and soil organic matter dynamics for the grassland biome worldwide [J]. Global Biogeochemical Cycles, 7: 785-890. |
| [26] | Potter C S, Randerson J T, Field C B, et al. 1993. Terrestrial ecosystem production: A process model based on global satellite and surface data [J]. Global Biogeochemical Cycles, 7: 811-841. |
| [27] | Prince S D, Goward S N. 1995. Global primary production: A remote sensing approach [J]. Journal of Biogeography, 22: 815-835. |
| [28] | 朴世龙, 方精云, 郭庆华. 2001. 1982~1999 年我国植被净第一性生产力及其时空变化[J]. 北京大学学报 (自然科学版), 37 (4): 563-569. Piao Shilong, Fang Jingyun, Guo Qinghua. 2001. Terrestrial net primary production and its spatio-temporal patterns in China during 1982-1999 [J]. Acta Scientiarum Naturalium Universitatis Pekinensis (in Chinese), 37 (4): 563-569. |
| [29] | Qian T T, Dai A G, Trenberth K E, et al. 2006. Simulation of global land surface conditions from 1948 to 2004: Part I: Forcing data and evaluations [J]. Journal of Hydrometeorology, 7: 953-975. |
| [30] | 冉有华, 李新, 卢玲. 2009. 四种常用的全球1 km土地覆盖数据中国区域的精度评价 [J]. 冰川冻土, 31 (3): 490-500. Ran Youhua, Li Xin, Lu Ling. 2009. Accuracy evaluation of the four remote sensing based land cover products over China [J]. Journal of Glaciology and Geocryology (in Chinese), 31 (3): 490-500. |
| [31] | Ran Y H, Li X, Lu L, et al. 2012. Large-scale land cover mapping with the integration of multi-source information based on the dempster-shafer theory [J]. International Journal of Geographical Information Science, 26 (1): 169-191. |
| [32] | Running S W, Hunt E R. 1993. Generalization of a forest ecosystem process model for other biomes, BIOME-BGC, and an application for global-scale models [C]./ Ehleringer J R. Scaling Physiological Processes: Leaf to Globe. San Diego, CA: Academic Press, 141-158. |
| [33] | Sitch S, Smith B, Prentice I C, et al. 2003. Evaluation of ecosystem dynamics, plant geography and terrestrial carbon cycling in the LPJ dynamic global vegetation model [J]. Global Change Biology, 9 (2): 161-185. |
| [34] | 陶波, 李克让, 邵雪梅, 等. 2003. 中国陆地净初级生产力时空特征模拟[J]. 地理学报, 58 (3): 372-380. Tao Bo, Li Kerang, Shao Xuemei, et al. 2003. Temporal and spatial pattern of net primary production of terrestrial ecosystems in China [J]. Acta Geographica Sinica (in Chinese), 58 (3): 372-380. |
| [35] | Uchijima Z, Seino H. 1985. Agroclimatic evaluation of net primary productivity of natural vegetations I. Chikugo model for evaluating net primary productivity [J]. Journal of Agricultural Meteorology, 40: 343- 353. |
| [36] | 王鹤松, 贾根锁, 冯锦明, 等. 2010. 我国北方地区植被总初级生产力的空间分布与季节变化 [J]. 大气科学, 34 (5): 882-890. Wang Hesong, Jia Gensuo, Feng Jinming, et al. 2010. Spatial distribution and seasonality of gross primary production in northern China [J]. Chinese Journal of Atmospheric Sciences (in Chinese), 34 (5): 882-890. |
| [37] | Wang Y P, Law R M, Pak B. 2010. A global model of carbon, nitrogen and phosphorus cycles for the terrestrial biosphere [J]. Biogeosciences, 7: 2261-2282. |
| [38] | Yuan W P, Liu S G, Zhou G S, et al. 2007. Deriving a light use efficiency model from eddy covariance flux data for predicting daily gross primary production across biomes [J]. Agricultural and Forest Meteorology, 143: 189-207. |
| [39] | Zaehle S, Sitch S, Smith B, et al. 2005. Effects of parameter uncertainties on the modeling of terrestrial biosphere dynamics [J]. Global Biogeochemical Cycles, 19 (3): doi: 10.1029/2004GB002395. |
| [40] | Zhao M S, Heinsch F A, Nemani R R, et al. 2005. Improvements of the MODIS terrestrial gross and net primary production global data set [J]. Remote Sensing of Environment, 95: 164-176. |
| [41] | 朱文泉, 陈云浩, 徐丹, 等. 2005. 陆地植被净初级生产力计算模型研究进展 [J]. 生态学杂志, 24 (3): 296-300. Zhu Wenquan, Chen Yunhao, Xu Dan, et al. 2005. Advances in terrestrial net primary productivity (NPP) estimation models [J]. Chinese Journal of Ecology (in Chinese), 24 (3): 296-300. |
| [42] | 朱文泉, 潘耀忠, 阳小琼, 等. 2007. 气候变化对中国陆地植被净初级生产力的影响分析 [J]. 科学通报, 52 (21): 2535-2541. Zhu Wenquan, Pan Yaozhong, Yang Xiaoqiong, et al. 2007. Analysis of the effects of climate change on net primary production of terrestrial vegetation in China [J]. Chinese Science Bulletin (in Chinese), 52 (21): 2535-2541. |